
Оригинал доступен на сайте tolweb.org
Стегоцефалы (панцирноголовые): четвероногие и другие позвоночные с пальцами
Мишель Лоран

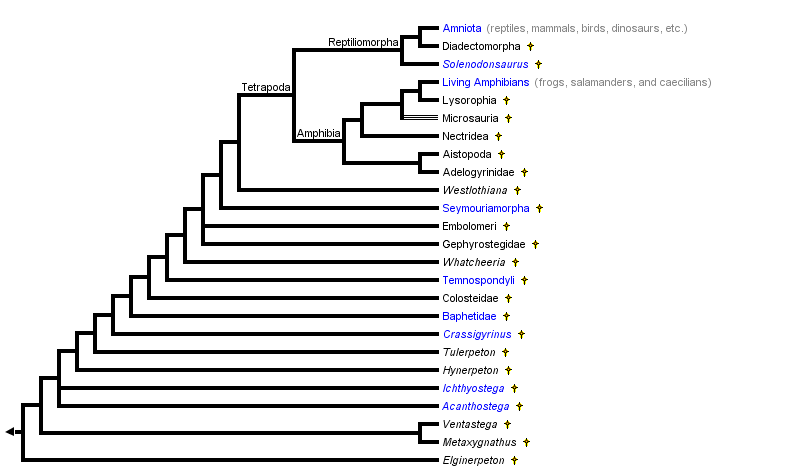
Модифицировано из Кэрролл (1995), Лоран и Райз (1997) и Лоран (1998a-c). Разряд Whatcheeria соответствует Ломбарду и Болту (1995). Разряд малоизвестных девонских таксонов (тех, количество пальцев которых является неопределенным: Elginerpeton, Metaxygnathus, Ventastega и Hynerpeton) соответствует Ahlberg (1998). Темноспондильные часто считаются ранними земноводными. Однако, обратите внимание, что в данном филогенезе они не являются тесно связанными с беспанцирными.
Содержащая группа: Лопастеперые рыбы
Вступление
На этой странице рассматриваются наземные позвоночные и другие позвоночные, у которых на конечностях присутствуют пальцы, а не плавники. Крепкая конечность, характерная для этой клады, обычно имеет четко выраженные суставы и пальцы (пальцы рук и ног) и носит название chiridium. Эта группа включает в себя около 21 100 существующих видов и, возможно, гораздо большее количество вымерших видов. Четвероногие — это только одна из нескольких групп позвоночных с chiridium (см. Раздел о классификации ниже).
Наземные позвоночные распространены по всему миру. Самые ранние представители этой группы были умеренно большими (с длиной тела 1-2,5 м). Самые древние из известных скелетных останков наземных позвоночных были обнаружены в верхнем девоне в Восточной Гренландии (Клак, 1994). Наличие троп в нижнем и среднем девоне в Австралии привело к предположению, что данная группа, возможно, возникла в период нижнего девона, по крайней мере, 400 000 000 лет назад (Уорен и др., 1986), но пальцы у этих оттисков не видны, поэтому указанные тропы могли быть оставлены другими лопастеперыми.
Самая большая группа наземных позвоночных — это Tetrapoda (см. Раздел «Классификация наземных позвоночных» ниже). «Tetrapoda» означает «четыре лапы», и группа была названа так, поскольку у ее представителей попросту были четыре конечности, а не плавники. Данный таксон включает в себя около 3 000 существующих видов амфибий (лягушек, саламандр и цецилий) и приблизительно 18 100 существующих видов амниот (млекопитающих, рептилий и птиц). Количество вымерших видов четвероногих, конечно, неизвестно, но около половины известных в настоящее время видов четвероногих вымерли (Кэрролл, 1988).
Четвероногие возникли не позднее миссисипского периода (около 350 миллионов лет назад) — периода, благодаря которому мы имеем представление о самых старых известных сородичах живых амфибий. Сородичи амниот, должно быть, существовали в то время, но они до сих пор не были найдены. Окаменелость, о которой речь велась ранее (Смитсон и др, 1994; Кэрролл, 1995) как о раннем миссисипском амниоте или антракозавре (Westlothiana), является, вероятно, либо stem-tetrapod, либо ранней амфибией (Лоран, 1998a). Стегоцефалы (определение этой группы см. в разделе о классификации ниже) возникли не позднее верхнего девона.
Общая длина четвероногих составляет от 9,8 мм (лягушка Psyllophryne didactyla) до 30 м (синий кит). Они распространены по всему миру и населяют все основные места обитания. Большинство из них наземные, но некоторые вернулись в водную среду, в которой жили наши далекие предки. К водным четвероногим относятся различные саламандры (sirenidae, cryptobranchidae, proteidae и т. д.), Лягушки (pipidae), некоторые цецилии (typhlonectidae), кожистые черепахи, морские змеи, ластоногие (тюлени и моржи) и киты. У некоторых четвероногих имеются способности к полету (птицы и летучие мыши), в то время как другие планируют (белки-летяги; Dermotherium, иногда называемые «летающими лемурами», даже при том, что они не приматы), и летающие драконы (Draco volans).
Страница История жизни стегоцефалов содержит информацию по этой непростой теме.
Основным органом дыхания большинства стегоцефалов является легкое, но у многих групп существуют и другие органы дыхания. Более подробная информация доступна на странице Дыхание у стегоцефалов.
У многих стегоцефалов имеется барабанная перепонка для того, чтобы слышать высокочастотные звуки, передающиеся по воздуху, а орган боковой линии обнаружен у многих водных амфибий. Для получения дополнительной информации посетите страницу Слух у стегоцефалов.
Характеристики
Стегоцефалы обладают обширной историей окаменелостей (Кэрролл, 1988). Филогенетические исследования выявили несколько выявленных характеристик (синапоморфий) у стегоцефалов:
- Исчезновение у представителей кое-каких костей черепа. У пандерихтисов (группы лопастеперых рыб, наиболее тесно связанных с панцерноголовыми), череп объединен несколькими костями с плечевым поясом очень крепко: указанные кости перестали существовать в самом начале эволюции наземных позвоночных. Исчезновение нескольких костей черепа также позволила создать подвижную шею, позволяющую двигать головой в отношении к туловищу. Такая особенность позволяет при ходьбе довольно статично держать голову.
- Исчезновение глазных костей, покрывающих жаберную камеру у костных рыб. Ранние хоановые не нуждаются в жаберной крышке, потому что внутренние жабры у них перестали существовать, которые присущи их ранним предкам. Тем не менее, жаберная оболочка исчезла, вероятно, еще до исчезновения внутренних жабр (Коутс и Клак, 1991).
- Изменения хорды и ригидного позвоночника. Тело позвоночника osteolepiforms является тонким и окружает хорду (жесткий стержень, присутствующий у всех хордовых и сохраняющийся у человека в качестве межпозвоночных дисков), сильно ee не сжимая. У стегоцефалов центральные слои толстые и сужают хорду. Специальные поверхности суставов (зигапофизы) соединяют нервные дуги друг с другом. Хорда короче, то есть такая, что не распространяется на черепную коробку. Хорда osteolepiforms распространялась до места, вблизи гипофиза.
- Четыре крепкие конечности с отдельными пальцами (пальцами рук и ног). У ранних лопастеперых рыб, снабженных плавниками, таких как Eusthenopteron, были крепкие плавники с элементами, гомологичными плечевой кости, лучевой кости, локтевой, связующего звена, локтевой, бедренной, большеберцовой, малоберцовой и пяточной костей, но гомологичность более дистальных элементов конечностей является неопределенной и спорной. В настоящее время неясно, являются ли сегментированные, разветвленные, эндоскелетные лучевые кости в дистальной части плавника лопастеперых, таких как Eusthenopteron или Tiktaalik, гомологичными пальцам. Были представлены как аргументы за (Шубин и др., 2006), так и аргументы против (Лоран, 2006) данной гипотезы.
- Крестцовое ребро, соединяющее осевой скелет (позвоночник) с тазовым поясом (бедром). Это позволяет передавать вес тела четвероногих на задние конечности. У osteolepiformes не существовало какой-либо костной связи между их осевым скелетом и тазовым поясом. Исчезновение лучей дермального плавника (модифицированные чешуйки, поддерживаеміе плавниками). Это просто означает удаление системы, в которой больше нет надобности и, вероятно, которая на суше будет даже мешать.
Указанные особенности появились не все сразу и не внезапно. У самые древних известных четвероногих, таких как Ichthyostega и Acanthostega, есть промежуточные параметры некоторых из этих особенностей и имеется недостаток в других. Например, Ichthyostega сохранил субоперкулярную кость — ту, которая составляла часть глазного комплекса, покрывавшего у osteolepiformes жаберную камеру. Acanthostega сохранил аноклетрум, являющийся одной из составляющих, связывавших череп с плечевым поясом у остеолипоформ (Коутс и Клак, 1991). Хорда у Ichthyostega и Acanthostega глубоко проникала в мозг, и у большинства ее хвостовых позвонков отсутствовали зигапофизы (Ярвик, 1952). Связь тазового пояса с крестцовым ребром у Acanthostega была еще плохо выражена. Наконец, как у Ichthyostega, так и у Acanthostega сохраняются лепидотрихии в хвосте, что указывает на то, что у упомянутых таксонов все еще был хвостовой плавник.
Предыдущий список включает исключительно характеристики скелета, потому что все самые ранние группы четвероногих вымерли, а пластичные анатомические части можно изучать только на примере представителей существующих таксонов. Следующие характеристики встречаются у четвероногих, а вот у других существующих позвоночных — нет. Это:
- Слой мертвых, роговых клеток, который уменьшает потери воды при испарении. Такой слой есть у амниот и у большинства беспанцирных. Кератин помогает поддерживать слои липидов, уменьшающих потери воды (сам кератин обладает плохими водонепроницаемыми свойствами).
- Мышечный язык с железами развит в значительной степени. И все же, у некоторых беспанцирных присутствует только основной язык, как у рыб. Основной язык — это просто мясистая складка на дне рта, где собственные мышцы отсутствуют, а подвижность ограничена.
- Паращитовидная железа участвует в контроле уровня кальция в крови.
- Гардерова железа, расположенная перед глазом. Эта железа выделяет маслянистую жидкость, смазывающую глаза.
- Вомероназальный орган (орган Якобсона). Этот орган обоняния расположен в небе и, вероятно, используется для того, чтобы чувствовать запах пищи, находящейся во рту.
- Потеря внутренних жабр. Внешние жабры, присутствующие у многих присобленных к воде беспанцирных и беспанцирных в стадии личинки, являются новыми структурами и не гомологичны внутренним жабрам рыб.
Трудно определить, когда именно появились указанные характеристики, потому что в окаменелостях они не сохранились (за исключением косвенных признаков внутренних жабр), а самые близкие известные родственники тетраподов вымерли. Однако эти признаки не обнаружены у двоякодышащих (ближайших сохранившихся родственников четвероногих). У Acanthostega — девонских стегоцефалов — все еще существовали внутренние жабры (Коутс и Клак, 1991), и Ichthyostega, вероятно, сохранил их тоже, но ни у каких других стегоцефалов, насколько известно, их не было. Следовательно, внутренние жабры, вероятно, были утрачены в начале эволюции стегоцефалов, в девоне или в миссисипском периоде (около 360 миллионов лет назад), и ни у одного четвероногого не было внутренних жабр.
Классификация наземных позвоночных
В прошлом большинство наземных хоановых были включены в надкласс четвероногих (Гафни, 1979). Недавно четвероногие были формально определены как краун-группа (Готье и др., 1989). Краун-группа — это клад, который включает последнего общего предка двух или более существующих таксонов и всех его потомков. В этом случае четвероногие была определена как клад, который включает в себя последнего общего предка беспанцирных и амниот, а также всех его потомков.
Согласно Готье и др. (1989), класс четвероногих включал большинство известных ископаемых наземных позвоночных, поскольку считается, что темноспондилы являются стем-группой амфибий, тогда как эмболомеры, гефиростегиды и сеймуриаморфы считались более тесно связанными с амниотами, чем с беспанцирными. Поэтому только несколько очень ранних наземных позвоночных, таких как Ichthyostega и Acanthostega, были исключены из класса четвероногих.
Филогенез хоановых, представленный здесь, предполагает, что темноспондилы, эмболомеры, гефиростегиды и сеймуриаморфы не являются частью краун-группы. Если это так, то указанные таксоны не относятся к четвероногим, и появление «конечности четвероногих» предшествует появлению самих четвероногих. В первых версиях этой страницы всех саркоптеригов, обладающих пальцами, называли просто наземными позвоночными, потому что формальная филогенетическая таксономия всей этой клады не была предложена. Указанная таксономия была опубликована недавно (Лоран, 1998a). Таксон стегоцефалов (который в течение длительного времени включал всех позвоночных, за исключением беспанцирных и амниот, имеющих chiridium), определял всех хоантовых как таких, что более тесно связанны с Темноспондилами, чем с Пандерихтисами (ближайшим сородичем четвероногих, о которых известно, что у них были парные плавники). Поэтому стегоцефалы включают в себя все таксоны, у которых есть пальцы, и несколько (Elginerpeton, Metaxygnathus, Ventastega и Hynerpeton), у которые могут обладать парными плавниками. Вопреки предыдущему использованию данного термина, стегоцефалами теперь называется кладой. Термин «панцирноголовые» будет использоваться ниже, потому что он не передает гипотетическую и, вероятно, несколько ошибочную интерпретацию того, что все позвоночные, имеющие пальцы, являются наземными. Как объясняется ниже, самые ранние представители этой клады, вероятно, были просто приспособлены к жизни в воде.
Примечание об узле, обозначенном как Amphibia: иные ограничивают название Amphibia потомками самого последнего общего предка существующих амфибий (терминальный таксон Живые Амфибии в этом дереве). Автор предпочитает использование, указанное в дереве, указанном выше, а определение амфибий как и всех четвероногих, связанных более тесно с существующими амфибиями, чем с амниотами, имеет исторический приоритет (Лоран, 1998a), но второе обозначение было довольно широко распространено и не может быть проигнорировано. На этой странице термин «Amphibia» всегда относится к узлу, указанному в дереве выше, а существующие амфибии называются беспанцирными.
Примечание об узле, обозначенном как Tetrapoda: другие расширяют использование указанного названия, чтобы включить всех позвоночных, имеющих конечности с пальцами (Лоран и Андерсон, 2004). Однако на этой странице Tetrapoda (четвероногие) всегда имеет в виду узел, указанный в дереве, расположенном выше.
Обсуждение филогенетических связей
Филогенез панцирноголовых является спорным. Почти все таксономисты сходятся во мнении, что существующие амфибии образуют монофилетическую группу (Lissamphibia — беспанцирные), которая тесно связана с амниотами, но происхождение беспанцирных и амниот является спорным. Лоран и Райз (1997, 1999), Лоран (1998a, b), а также Уаллин и Лоран (2004) предполагают, что тонкопозвонковые являются стем-группами амфибий и что диадэктоморфы более тесно связаны с амниотами, чем с беспанцирными. Следовательно, многие группы палеозойских панцирноголовых, такие как темноспондилы и сеймоуриаморфы, с амниотами или беспанцирными тесно не связаны:

Тем не менее, в самых ранних исследованиях и в нескольких недавних исследованиях предполагалось, что беспанцирные происходят от темноспондилов и что диадэктоморфы, сеймуриаморфы и эмболомеры более тесно связаны с амниотами, чем с беспанцирными (Болт, 1969; Гафни, 1979; Готье и др., 1988, 1989; Милнер, 1988, 1993; Панчен и Смитсон, 1988; Труб и Клотье, 1991; Ломбард и Сумида, 1992; Рута и др., 2003а, б; Рута и Коутс, 2007). Тонкопозвонковые часто рассматривались как парафилетическая группа, в которую входили ранние сородичи темноспондилов и беспанцирных. Поэтому большинство известных стегоцефалов рассматривались как связанные с беспанцирными или амниотами. Считалось, что только Ichthyostega и другие девонские таксоны (Acanthostega, Tulerpeton) принадлежат к одной из этих двух основных эволюционных линий:

Некоторые палеонтологи все еще предлагают полифилетическое происхождение существующих амфибий (Андерсон, 2007, 2008; Кэрролл, 2007; Андерсон и др., 2008), где безногие земноводные относятся к тонкопозвонковым, а безхвостые и хвостатые земневодные — к темноспондилам. Это подразумевает, что гимнастофионаны более тесно связаны с амниотами, чем с бесхвостыми и хвостатыми земноводными. Данный филогенез несовместим со всеми опубликованными молекулярными и наиболее морфологическими филогенезами, предполагающими, что существующие амфибии образуют кладу, в которой амниоты исключены (Лоран, 2002). Кроме того, пересмотр шифрования основной поддерживающей матрицы показывает, что она фактически поддерживает монофилетическое происхождение беспанцирных из «тонкопозвонковых» (Марьянович и Лоран, 2009). Данные о развитии также были предложены в поддержку существующей полифилии амфибий (Шох и Кэрролл, 2003; Фрьобиш и др., 2007), но эти аргументы были опровергнуты (Шох, 2006; Жермен и Лоран, 2009; Марьянович и Лоран, 2009). Данные о развитии не проводят различий между разными гипотезами о происхождении беспанцирных.
Самый недавний филогенез, отстаивающий полифилию беспанцирных (Андерсон и др., 2008), частично основан на недавно описанном temnospondyl Gerobatrachus hottoni, который первоначально интерпретировался как сестринская группа Batrachia, которая гнездится глубоко в темноспондилах; безногие земневодные относятся к тонкопозвонковым. Этот вывод частично основан на интерпретации различных морфологических признаков рода Gerobatrachus, таких как ножка зубов (синапоморфия беспанцирных), нахоящаяся в основе зуба (слитые дистальные лапки 1 и 2; синапоморфия земноводных) и узкая, направленная вбок неба (синапоморфия salientia). Тем не менее, наличие синапоморфий нескольких клад, а также другие, гораздо более простых признаков, таких как сохранение постпариетальности, пластиначтости и супратемпоральности, указывает на обширную гомоплазию. Возможны другие интерпретации морфологии Gerobatrachus hottoni, и включение этого вида в первую матрицу данных, которая поддерживает полифилию беспанцирных (Макгоуен, 2002), все еще поддерживает монофилетическое происхождение беспанцирных в границах тонкопозвонковых (Марьянович и Лоран, 2008b).
В нескольких молекулярных исследованиях были предложены очень ранние даты начала диверсификации беспанцирных, начиная примерно с 337 млн лет назад (Чжан и др., 2005) до 362 млн лет назад (Сан Мауро и др., 2005; Ролантс и др., 2007). , Чжан и др. (2005) также предположили, что это подразумевало, что беспанцирные появились от темноспондилов, но Ли и Андерсон (2006) указали, что такие ранние даты диверсификации более совместимы с полифилетической гипотезой, согласно которой существующие амфибии образуют кладу, которая исключает амниотов, но происходит от тонкопозвонковых, и темноспондил. Тем не менее, новое молекулярное и палеонтологическое датирование диверсификации беспанцирных предполагает гораздо более недавнюю диверсификацию для этой группы, в перми (Марьянович и Лоран, 2007). Расчет доверительного диапазона истинного стратиграфического периода беспанцирных, основанный на дальнейшей разработке самого сложного метода, разработанного Маршаллом (1997), также предполагает пермское монофилетическое происхождение группы (Марьянович и Лоран, 2008a).
Дополнительное обсуждение и более подробную информацию о филогенезе можно найти на странице Филогенез стегоцефалов.
Происхождение стегоцефалов
Было предложено несколько сценариев, объясняющих происхождение chiridium и причины, по которым позвоночные выходят на сушу. Теория, найденная в большинстве популярных книг, гласит, что засушливый климат, который когда-то считался преобладающим в девоне, заставил наших лопастеперых предков выползти из водоемов (которые в определенный сезон высыхали), чтобы достичь более крупных и глубоких водоемов (Ромер, 1933). Однако проблема с этим сценарием заключается в том, что девон больше не считается таким периодом, когда были сезонные засухи, а классические позднедевонские образования, такие как эскуминак в Квебеке, Канада, в настоящее время считаются устьичными (следовательно, сезонная засуха не может возникнуть, так как уровень моря в значительной степени не зависит от времени года). Эндоскелет плавников тетраподоморфов, у которых наличествовали плавники — недавно изученный у семейства трихоптеридов из рода Eusthenopteron — был, по-видимому, очень губчатым и, по-видимому, не подходил для того, чтобы выдерживать значительные механические нагрузки в течение длительных периодов времени, как это происходило бы при длительных наземных прогулках (Лоран и др., 2007). Кроме того, недавние исследования функциональной морфологии выдвигают предположение, что конечности могли развиваться, чтобы позволить нашим ранним предкам ходить по дну мелких водоемов или болот (Эдвардс, 1989). Открытие довольно исчерпывающих образцов девонских хоанов Acanthostega (Коутс и Кларк, 1990) подтверждает, что их конечности были плохо приспособлены для прогулок по суше (Циммер, 1995). Предполагаемое наличие внутренних жабр у Acanthostega также подтверждает, что это животное все еще было в основном водным (Коутс и Кларк, 1991). Переход от водной среды к земной, похоже, занял больше времени, чем считалось ранее.
Ранняя эволюция конечностей
Давно известно, что компоненты, гомологичные стилоподию (бедро и плечевая кость) и зевоподию (лучевая кость, локтевая кость, большеберцовая кость и малоберцовая кость), встречаются у тетроподоморфов (как говорится, например, у Грегори, 1911), но до недавних пор мало что было известно о времени появления различных признаков конечностей у четвероногих. В последнее время на этом фронте достигнут большой прогресс. Например, недавнее гистологическое исследование показало, что парные плавники Eusthenopteron foordi обладали настоящими длинными костями, которые росли в длину и толщину благодаря эндохондральному и периостальному окостенению, как и у четвероногих (Менье и Лоран, в печати). Бойсверт и др. (2008) показали, что дистальная часть грудного плавника у Panderichthys (одного из тетраподоморфов, снабженных плавниками, наиболее близких к краун-группе) состоит из нескольких радиалов, как и следовало ожидать от вида, тесно связанного с панцироголовыми. Более ранняя реконструкция, которая представляла одну большую дистальную пластинку вместо нескольких меньших радиальных линий, представляется неверной интерпретацией.
До недавнего времени самым древним неопровержимым доказательством наличия пальцев были остатки скелета с самого конца девона (фаменский период). Четыре девонских таксона, согласно предположениям включенных в дерево, представленное выше, известны из фрагментарных останков. Фрагментарная природа останков этих родов не позволяет достоверно оценить их родство, хотя Коутс (1996) и Альберг (1998) предположили, что они более тесно связаны с четвероногими, чем с родом panderichthys (ближайшими известными родственниками четвероногих, которые, как известно, сохранили парные плавники). К таким загадочным таксонам относятся Elginerpeton (Альберг, 1995), Ventastega (Альберг и др., 1994), Metaxygnathus (Кэмпбелл и Белл, 1977) и Hynerpeton (Дейшлер и др., 1994). Их аппендикулярный скелет плохо известен (или не известен), и они исключены из наименьшей клады, в которую входят позвоночные, имеющие конечности. Следовательно, они могли сохранять плавники (Лоран и др, 2000). Однако недавнее обнаружение тропы относящейся к периоду среднего девона (Eifelian) (Нидзвецкий и др., 2010) в заброшенном карьере Захельме (Польша) повышает вероятность того, что такие формы, как Elginerpeton, обладают пальцами. Далее Нидзвецкий и др. (2010) предполагают, что эта тропа подразумевает, что у девонских стегоцефалов очень плохая окаменелость, поскольку это предполагает (с их точки зрения) наличие шести длинных призрачных линий, простирающихся от Среднего до позднего девона у стегоцефалов, кроме того, к дополнительным линиям привидений у близкородственных тетраподоморфов, таких как Panderichthy, Tiktaalik, Elpistostege и Livonia (см. рисунок, часть A). Тем не менее, данное утверждение может быть преувеличено в силу того, что точное систематическое положение создателя тропы в Захельмье неизвестно. Он может представлять более базальную группу, исключенную из наименьшей клады, которая включает в себя Elginerpeton, Ventastega и более краун-групповые виды, а это будет означать одну безымянную родственную стегоцефалам линию, простирающуюся от середины до позднего девона, в дополнение к нескольким безымянным линиям тетраморфов, обладающих плавниками (Лоран, 2010; рисунок, часть B).
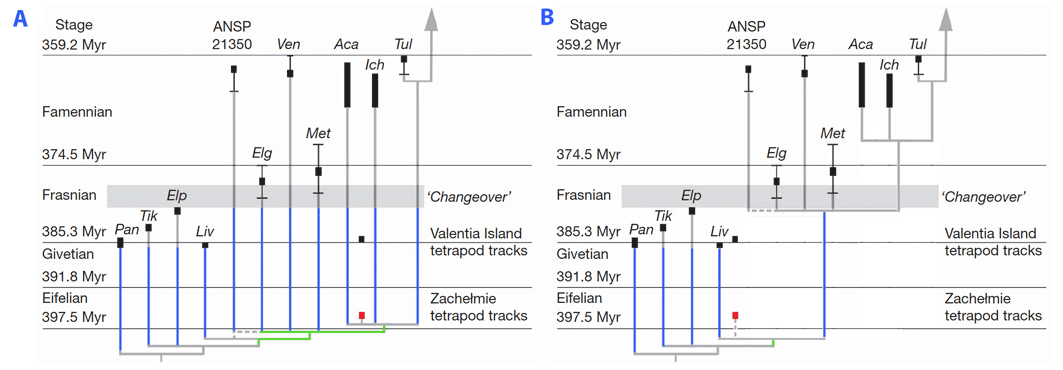
Возможные филогенетические расположения польской среднедевонской тропы. А — расположение, предложенное Нидзвецким и др. (2010). B — расположение, предложенное (в тексте) Лораном (2010). Учитывая имеющиеся доказательства, ни одно из них не может быть опровергнуто. Безымянные роды обозначены синим цветом. Ветви чуть ниже возможного положения тропы обозначены зеленым цветом. Часть А перенесена с рисунка 5b Нидзвецким и др. (2010).
Считается, что наличие пяти пальцев на передних и задних конечностях у четвероногих является зачаточным и это самое большое число пальцев, которое было у наших ранние предки. Пять пальцев, вероятно, были в наличии у самых ранних четвероногих, но не у самых ранних панцироголовых. Недавние открытия показали, что у самых ранних стегоцефалов было больше пальцев; У Acanthostega было восемь пальцев на передних конечностях, у Ichthyostega было семь пальцев на задних, а у Tulerpeton было шесть пальцев на передних (Лебедев, 1986; Гулд, 1991). Однако эти «лишние» пальцы были быстро утеряны, поскольку, как известно, ни у одного постдевонского панцероголового не было более пяти пальцев.
Коутс (1991, 1996) предположил, что пентадактильность возникала дважды (по крайней мере, в случае с передней конечностью), потому что в его предполагаемых филогенезах девонский полидактильный панцироголовый Tulerpeton тулерпетон, как полагается, был более тесно связан с амниотами, чем с беспанцирными. Поэтому он утверждал, что восстановление от полидактильного до пентадактильного состояния происходило у антракозавров (амниот и их вымерших родственников) и у амфибий (хотя пентадактильность у амфибий известно только исходя из нескольких образцов; у большинства беспанцирных есть только четыре пальца в кисти). Однако повторный анализ этих данных и включение Tulerpeton в матрицу данных, которая включает больше таксонов, позволяет предположить, что пентадактильность появилась только один раз, задолго до расхождения между амфибиями и антракозаврами (Лоран, 1998c). Этот вывод является результатом размещения Tulerpeton в более базальной позиции в филогенезе за пределами клады, включающем все пост-девонские (и все пентадактильные) таксоны.
Завоевание суши позвоночными
Первые позвоночные с конечностями, такие как Acanthostega и Ichthyostega, принадлежали к таксонам, явно обитающим в воде (Клак, 2002), потому что они, по-видимому, сохранили внутренние жабры, о чем свидетельствуют бороздки на кератобранхиальных мышцах, которые могли вместить афферентные жаберные артерии. Они также сохранили постбранхиальную пластинку лимана, которая могла отграничивать ветвистую камеру сзади (Коутс и Клак, 1991). Кроме того, они сохранили лепидотрихию в хвостовом плавнике и хорошо развитый орган боковой линии, как показывают каналы его головной части.
Внутренние жабры, возможно, исчезли довольно рано во времена каменноугольного периода, и у панцирноголовых нет никаких следов lepidotrichia после девона, и это может указывать на немного менее водный образ жизни. Тем не менее, орган боковой линии сохранился у большинства каменноугольных и многих пермских панцирноголовых , что говорит о том, что многие виды все еще были преимущественно водными. Отсутствие канавок для органа боковой линии не является надежным критерием для определения образа жизни, соответствующего суше, поскольку орган может присутствовать, не оставляя следов на скелете. Поскольку форма тела ранних панцирноголовых довольно консервативна, водные виды не обязательно сильно отличаются от наземных по форме тела (Лоран, 2008). Таким образом, дополнительные критерии, такие как микроанатомия кости, были изучены, чтобы определить, когда позвоночные стали наземными (Лоран и др., 2004; Жермен и Лоран, 2005). Предварительные результаты предполагают, что у панцирноголовых в раннем карбоне появился наземный образ жизни (Крылов и др., 2008), но для получения надежных результатов придется изучить еще много видов.
Концентрация кислорода в атмосфере, по-видимому, могла сыграть роль в том, чтобы позвоночные могли перемещаться на сушу. Первые водные панцирноголовые появились, когда атмосферный кислород присутствовал в низкой концентрации. Разрыв Ромера, период, в течение которого известно очень мало панцирноголовых, представляет собой заключительную фазу этого эпизода низкой концентрации кислорода. Вскоре после того, как концентрация кислорода в атмосфере достигла и даже превысила свой нынешний уровень, панцирноголовые подверглись впечатляющему эволюционному разветвлению, которое включало в себя первых наземных позвоночных. Это эволюционное разветвление параллельно происходило у членистоногих. Так, Вард и др. (2006) предполагают, что концентрация кислорода вызывала или облегчала деятельность членистоногих и позвоночных животных на суше, и что концентрация кислорода около 20% в атмосфере была нижним порогом, который позволил перейти от жаберного дыхания в воде к дыханию с использованием воздуха у позвоночных и членистоногих.
Первоначальная среда обитания стегоцефалов
Долгое время предполагалось, что первыми стегоцефалами были пресноводные виды, подобные существующим амфибиям и дипноанам (Ромер, 1933). Тем не менее, многие исследования, проведенные в 1980-х годах, показали, что ранние саркоптеригианы (включая дипноаны) чаще всего были морскими формами (Жанвьер, 1996). Аналогичным образом, недавно было показано, что несколько классических пермокарбононосных местностей, которые дали панцирноголовым и которые долгое время считались (по крайней мере, палеонтологами позвоночных) репрезентативными средами, проявляют морское влияние (Лоран и Солер-Гийон, 2006, 2010; Шульц, 2009). Представляется вероятным, что многие из самых ранних панцирноголовых были прибрежными формами, о чем свидетельствуют среднедевонские тропы, которые были оставлены в приливной или лагунной среде (Нидзевский и др. , 2010) и в явно морской среде обитания Тулерпертона (Лебедев, 1986). ). Однако некоторые их близкие родственники, такие как Tiktaalik (Дешлер и др., 2006) и некоторые ранние панцирноголовые, такие как Ichthyostega и Acanthostega (Клак, 2002), были интерпретированы как пресноводные формы. Вполне вероятно, что в позднем палеозое панцирноголовые обитали в соленой, пресной и наземной среде. Непереносимость соленой воды, которая характерна для большинства беспанцирных, по-видимому, является аутапоморфией беспанцирных (Лоран и Солер-Гийон, 2010).
Другие названия наземных позвоночных
Stegocephalians
Рекомендованная литература
Альберг П. Э. 1995. Elginerpeton pancheni и самая ранняя клада четвероногих. Nature, 373: стр. 420-425.
Альберг П. Э. 1998. Посткраниальный ствол, оставшийся у четвероногих после девона. Скат Крейг, Морайшир, Шотландия. Zoological Journal of the Linnean Society, 122: стр. 99-141.
Альберг П. Э., Э. Лукшевичс и О. Лебедев. 1994. Первые находки четвероногих из девона (верхнего фамеского яруса)в Латвии. Философские труды Королевского общества, B, 343: стр. 303-328.
Андерсон, Дж. С. 2007 Включение онтогенеза в матрицу: филогенетическая оценка доказательств развития касательно происхождения современных амфибий. В: Большой переход в эволюции позвоночных, (ред. Андерсона Дж. С и Сьюса Х.-Д.). Стр. 182-227. Indiana University Press, Блумингтон.
Андерсон Дж. С. 2008. Происхождение современных амфибий. Evolutionary Biology, 35: 231–247.
Андерсон Дж. С., Р. Р. Рейс, Д. Скотт, Н. Б. Фребиш и С. С. Сумида. 2008. Безпанцирные периода ранней перми из Техаса, а также происхождение лягушек и саламандр. Nature, 453: стр. 515–518.
Бойсверт С. А., Э. Марк-Курик и П. Э. Альберг. 2008. Грудной плавник Panderichthys и происхождение пальцев. Nature, 456: стр. 636–638.
Болт, Дж. Р. 1969. Происхождение беспанцирных: вероятные протобеспанцирные периода нижней перми из Оклахомы. Science, 166: стр. 888-891.
Кэмпбелл К. С. У. и М. У. Белл. 1977. Первобытная амфибия периода позднего девона из Нового Южного Уэльса. Alcheringa, 1: 369381.
Кэрролл, Р. Л. 1988. Палеонтология и эволюция позвоночных. Нью-Йорк: W. H. Freeman and Company.
Кэрролл Р. Л. 1995. Проблемы в филогенетическом анализе палеозойских хоановых. Бюллетень Национального музея естественной истории Парижа, 4-я серия 17: стр. 389-445.
Кэрролл Р. Л. 2007. Палеозойские предки саламандр, лягушек и цецилий. Zool. J. Linn. Soc. 150: стр. 1–140.
Клак, J.A. 1994. Черепа самых ранние из известных четвероногих, а также эволюция стремечка и овального окна. Nature, 369: стр. 392-394.
Клак Дж. А. 2002. Набирая обороты: происхождение и эволюция четвероногих. Блумингтон: Indiana University Press, 369 с.
Коутс М. 1991. Новый палеонтологический вклад в онтогенез и филогенез конечностей. В: Дж. Р. Хинчклифф (ред.) Пути развития конечностей позвоночных, стр. 325-337. Нью-Йорк: Plenum Press.
Коутс М. И. 1996. Девонские четвероногие Acanthostega gunnari; Ярвик: постчерепная анатомия, базальные взаимосвязи четвероногих и примеры эволюции скелета. Труды Королевского общества Эдинбурга, 87: стр. 363-421.
Коутс, М. И. и Дж. А. Клак. 1990. Полидактилия конечностей самых ранних известных четвероногих. Nature, 347: стр. 66-69.
Коутс, М. И. и Дж. А. Клак. 1991. Рыбоподобные жабры и дыхание у самого раннего из известных четвероногих. Nature, 352: стр. 234-236.
Дейшлер E. B., Н. Х. Шубин, К. С. Томсон и В. В. Амарал. 1994. Девонские четвероногие из Северной Америки. Science, 265: стр. 639-642.
Дейшлер E. B., Н. Х. Шубин и Ф. А. Дженкинс мл. 2006. Девонская рыба, похожая на четвероногих, и эволюция строения тела четвероногих. Nature, 440: стр. 757-763.
Эдварс, J. 1989. Две перспективы развития конечности четвероногих. The American Zoologist, 29: стр. 235-254.
Фрьобиш Н. Б., Р. Л. Кэрролл и Р. М. Шох. 2007. Окостенение конечностей в палеозойском у апатеон семейства branchiosauridae (Темноспондильные) и ранняя эволюция преаксиального доминирования в развитии конечностей четвероногих. Evolution & Development, 9: стр. 69-75.
Гафни Е. С. 1979. Монофилия четвероногих: филогенетический анализ. Бюллетень Музея естествознания им. Карнеги, 13: стр. 92-105.
Готье Дж., А. Г. Клюге и Т. Роу. 1988. Ранняя эволюция амниот. В М. Дж. Бентон (ред.) Филогенез и классификация четвероногих, Том 1: земноводные, рептилии, птицы: стр. 103-155. Oxford: Clarendon Press.
Готье Дж., Д. С. Каннателла, К. Де Кейроз, А. Г. Клюге и Т. Роу. 1989. Филопения четвероногих. В Б. Фернхольм, К. Бремер и Х. Йорнваль (ред.) The Hierarchy of Life: стр. 337-353. Нью-Йорк: Elsevier Science Publishers B.V. (Biomedical Division).
Жермен Д. и М. Лоран. 2005. Микроанатомия лучевой кости и образ жизни у амниот (позвоночные, четвероногие). Zoologica Scripta, 34: стр. 335-350.
Жермен Д. и М. Лоран. 2009. Эволюция ряда окостенений у саламандр и происхождения хвостатых земноводных, проанализированные с помощью синхронизации событий и не существовавших ранее методов. Evolution & Development, 11: стр. 170–190.
Гулд С. Дж. Восемь (или меньше) маленьких поросят. Natural History, январь 1991 года, стр. 22-29.
Грегори В. К. 1911. Конечности Эриопса и происхождение парных конечностей от плавников. Science 33: стр. 848-849.
Жанвьер P. 1996. Ранние позвоночные. Оксфордские монографии по геологии и геофизике. Оксфорд: Oxford University Press, 393 с.
Ярвик Э. 1952. О рыбоподобном хвосте у стегоцефалов семейства Ichthyostegidae с описаниями нового стегоцефала и нового кистеперого из верхнего девона восточной Гренландии. Meddelelser om Grønland, 114: стр. 1-90.
А. Крилов, Д. Жермен, А. Кановиль, П. Винсент, М. Саше и М. Лоран. 2008. Эволюция микроанатомии костей голеней четвероногих и применение ее в палеобиологическом заключении. Journal of Evolutionary Biology, 21: стр. 807–826.
Лоран М. 1998а. Важность глобальной парсимонии и исторического предубеждения в понимании эволюции четвероногих. Часть I — систематика, эволюция среднего уха и устройство челюстей. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: стр. 1-42.
Лоран М. 1998b. Важность глобальной парсимонии и исторического предубеждения в понимании эволюции четвероногих. Часть II — центр позвоночника, реберная вентиляция и педоморфоз. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série, 19: стр. 99-114.
Лоран М. 1998c. Переоценка происхождения пятиперстности. Evolution, 52: стр. 1476-1482.
Лоран, M. 2002. Филогения четвероногих, происхождение земноводных и определение наименования четвероногих. Syst. Biol., 51: стр. 364-369.
Лоран, M. 2006. Скудные свидетельства и изменчивые точки зрения на развивающийся аппендикс. Zoologica Scr. 35: стр. 667-668.
Laurin M. 2008. Systématique, paléontologie et biologie évolutive moderne : l’exemple de la sortie des eaux des vertébrés. Collection Parcours LMD – Sciences de la vie et de la Terre, ed. J. Segarra. Paris: Ellipses, 176 pp.
Лоран М. 2010. Как позвоночные покинули воду. Перевод М. Лорана. Беркли: University of California Press, XV + 199 с.
Лоран М. и Дж. С. Андерсон. 2004. Значение наименования четвероногих в научной литературе: обмен. Systematic Biology, 53: стр. 68-80.
Лоран, M. и Р. Р. Райз. 1997. Новый взгляд на филогенез четвероногих. В С. Сумида и К. Мартин (ред.) Происхождение амниот: завершение перехода на сушу: стр. 9-59. Лондон: Academic Press.
Лоран M. и Р. Р. Райз. 1999. Новое исследование Solenodonsaurus janenschi и пересмотр происхождения амниот и эволюции стегоцефалов. Canadian Journal of Earth Sciences, 36: стр. 1239-1255.
Лоран, M., Жирондот, M. и де Рикле, A. 2000. Ранняя эволюция четвероногих. Trends Ecol. Evol., 15: стр. 118-123.
Лоран М., Жирондот М. и М.-М. Лот. 2004. Эволюция микроанатомии длинных костей и образа жизни у беспанцирных. Paleobiology, 30: стр. 589-613.
Лоран М. и Р. Солер-Гийон. 2006. Самый древний из известных стегоцефалов (Sarcopterygii: Temnospondyli) из Испании. Journal of Vertebrate Paleontology, 26: стр. 284-299.
Лоран М. и Р. Солер-Гийон. 2010. Осмотическое отклонение и среда обитания ранних стегоцефалов: косвенные данные из парсимонии, тафономии, палеобиогеографии, физиологии и морфологии. В кн .: М. Веколи и Дж. Климента (ред.). Процесс заболачивания: моделирование сложных взаимодействий на границе биосферы и геосферы (339): стр. 151–179. Лондон: Геологическое общество Лондона.
Лебедев О. А. 1986. Первое упоминание о девонском четвероногом в СССР. Доклады: Раздел наук о Земле, 278: стр. 220-222.
Ли М.С. И. и Андерсон Дж. С. 2006. Молекулярные часы и происхождение современных амфибий. Mol. Phyl. Evol. 40: стр. 635-639.
Ломбард, Р. Э. и Дж. Р. Болт. 1995. Неизвестное ранее примитивное четвероногое Whatcheeria deltae раннего карбонного периода из Айовы. Palaeontology, 38: стр. 471-494.
Ломбард, Р. Э. и С. С. Сумида. 1992. Недавний прогресс в понимании ранних четвероногих. The American Zoologist, 32: стр. 609-622.
Марьянович, д. & Лоран М. 2007. Окаменелости, молекулы, времена расхождения и происхождение беспанцирных. Syst. Biol., 56: стр. 369-388.
Марьянович Д. и М. Лоран. 2008a. Оценка доверительных диапазонов стратиграфических подразделений высших таксонов: ситуация с Lissamphibia. Acta Palaeontologica Polonica, 53: стр. 413–432.
Марьянович Д. и М. Лоран. 2008b. Переоценка доказательств, подтверждающих неортодоксальную гипотезу о происхождении доживших до наших дней амфибий. Contributions to Zoology, 77: стр. 149–199.
Марьянович Д. и М. Лоран. 2009. Происхождение современных амфибий: комментарий. Evolutionary Biology, 36: стр. 336–338.
Маршалл С. Р. 1997. Доверительные диапазоны стратиграфических подразделений с неслучайным распределением горизонтов с ископаемыми. Paleobiology, 23: стр. 165-173.
Макгоуен, G.J., 2002.Земневодные Albanerpetontid из нижнего мела Испании и Италии: описание и пересмотр их систематики. Zool. J. Linn. Soc., 135: стр. 1-32.
Менье Ф. Ж. и М. Лоран в прессе. Микроанатомическое и гистологическое исследование длинных костей девонской лопастеперой рыбы Eusthenopteron foordi. Acta Zoologica 19 стр., 3 рисунка, 1 таблица (ранняя версия доступна на веб-сайте журнала).
Мильнер А. Р. 1988. Взаимосвязь и происхождение живых амфибий. В М. Дж. Бентон (ред.) Филогения и классификация четвероногих, Том 1: Амфибии, Рептилии, Птицы, стр. 59-102. Oxford: Clarendon Press.
Мильнер, А. Р. 1993. Палеозойские сородичи беспанцирных. Герпетологические монографии 7: стр. 8-27.
Нидзвецкий Г., П. Шрек, К. Наркевич, М. Наркевич, П. Э. Альберг, 2010. Тропы четвероногих из периода раннего Среднего Девона в Польше. Nature, 463: стр. 43–48.
Нуссбаум, Р. А. 1977. Rhinatrematidae: новое семейство цецилий (Amphibia: Gymnophiona). Периодические доклады Музея зоологии Мичиганского университета, 682: стр. 1-30.
Панчен, А. Л. и Т. Р. Смитсон. 1988. Взаимосвязи между самыми ранними четвероногими. В М. Дж. Бентон (ред.) Филогения и классификация четвероногих, том 1: земноводные, рептилии, птицы: стр. 1-32. Oxford: Clarendon Press.
Ролантс, К., Гауэр, Д. Дж., Уилкинсон М., Лоадер, С. П., Биджу, С. Д., Гийом К., Морио Л. и Боссайт Ф., 2007. Глобальные модели диверсификации в истории современных земноводных. Proc. Natl. Acad. Sci. U.S.A., 104: стр. 887-892.
Ромер А. С. 1933. Палеонтология позвоночных. 1-е изд., Чикаго: University of Chicago Press.
Рута, М. & Коутс, М. И. 2007. Периоды, узлы и конфликт характеристик: обращение к проблеме происхождения беспанцирных. J. Syst. Paleontol., 5: стр. 69-122.
Рута М., М. И. Коутс и Д. Д. Л. Квик. 2003. Еще раз о ранних взаимодействиях четвероногих. Биологические обзоры Кембриджского философского общества, 78: стр. 251-345.
Рута М., Дж. Э. Джеффри и М. И. Коутс. 2003. Супер-родословная ранних четвероногих. Труды Королевского общества Лондона, серия B, 270: стр. 2507-2516.
Сан Мауро Д., Венсес, М., Алькобендас, М., Зардоя, Р. и Мейер, A. 2005. Первоначальная диверсификация живых амфибий произошла раньше, чем распад Пангеи. Am. Nat., 165: стр. 590-599.
Шох Р.Р. 2006. Онтогенез черепа: паттерны развития рыб, сохраняющиеся по основным кладам четвероногих. Evolution & Development, 8: стр. 524-536.
Шох Р. Р. и Р. Л. Кэрролл. 2003. Онтогенетические доказательства палеозойского происхождения саламандр. Evolution & Development, 5: стр. 314-324.
Шульце Х.-П. 2009. Интерпретация морских и пресноводных палеосред в пермо-карбоновых отложениях. Палеогеография, Палеоклиматология, Палеоэкология, 281: стр. 126–136.
Шубин Н. Х., Дешлер Э. Б. и Дженкинс Ф. А. мл., 2006. Грудной плавник Tiktaalik roseae и происхождение конечности четвероногих. Nature, 440: стр. 764-771.
Смитсон, Т. Р., Р. Л. Кэрролл, А. Л. Панчен и С. М. Эндрюс. 1994. Westlothiana lizziae с Визейского о-ва Восточный Кирктон, Западный Лотиан, Шотландия и клады амниот. Труды Королевского общества Эдинбурга, 84: стр. 383-412.
Труб Л. и Р. Клотье. 1991. Филогенетическое исследование внешних и внутренних свзей Панцирных (Амфибии: Темноспондильные). В Х.-П. Шульце и Л. Труб (ред.) Происхождение высших групп четвероногих — Противоречия и консенсус: стр. 223-313. Итака: Comstock Publishing Associates.
Уаллин Г. и М. Лоран. 2004. Морфология черепа и сходство ряда Microbrachis, а также переоценка филогении и образа жизни первых земноводных. Journal of Vertebrate Paleontology, 24: стр. 56-72.
Уорд П., С. Лабандейра, М. Лоран и Р. Бернер, 2006. Подтверждение Разрыва Ромера как периода с низким уровнем кислорода, ограничивающего время начала наземной трансформации членистоногих и позвоночных. Слушания Национальной Академии Наук Соединенных Штатов Америки, 103: 16818-16822.
Уоррен, А., Р. Джапп и Б. Болтон, 1986. Самая первая тропа четвероногих. Alcheringa, 10: стр. 183-186.
Чжан П., Чжоу Х., Чен Ю.К., Лю Ю.Ф. и Кю, Л.-Х. 2005. Митогеномные точки зрения на происхождение и филогению живых амфибий. Syst. Biol., 54: стр. 391-400.
Циммер Ч., Выходя на сушу. Discover, июнь 1995 года, стр. 118-127.
Информация в интернете
Для общего ознакомления с четвероногими и их близкими родственниками, попробуйте Введение в тетраподов (UCMP, Berkeley).
Существует перевод этой страницы на Албанский язык, любезно предоставленный Радкой Марич, перевод на испанский от emfurn.com и перевод на малазийский от Линли Ло из Down To Five.
Правовладение иллюстрациями
 Научное название: Acanthostega gunnari Ichthyostega
Научное название: Acanthostega gunnari Ichthyostega
Комментарии: Девонские наземные хоановые Acanthostega (внизу) и Ichthyostega (сбоку).
Ссылки: впервые опубликована в журнале Discover
Создатель: Иллюстрация Альфреда Камаджяна
Статус образца — Ископаемое — Период: Девон
Copyright © 1995 Альфред Камаджян
Об этой странице
Я хотел бы поблагодарить г-на Джона Хатчинсона, г-жу Патрисию Лай и г-на Мэтью Марлоу, которые редактировали эту страницу. Я в долгу перед доктором Дэвидом Мэддисоном, который оказал неоценимую помощь в форматировании этой страницы, ее линковке с другими страницами на сайте Tree of Life, и чьи многочисленные предложения улучшили представление этой страницы. Я также благодарю Катю Шульц за проверку моих периодических обновлений перед их публикацией.
Национальный музей естествознания, Париж, Франция
Корреспонденцию по поводу этой страницы необходимо отправлять Мишелю Лорану по адресу [email protected]
Page Copyright © 2011Мишель Лоран
Добавить комментарий